
TRANSPORTE DE NUTRIENTES
Lo primero que tiene que hacer un microorganismo a la hora de su nutrición es captar los nutrientes que necesite desde el medio exterior. Debido a que la bicapapa lipídica actúa como barrera que impide el paso de la mayor parte de las sustancias, esto significa que deben existir mecanismos específicos para lograr la entrada de los nutrientes. Además, teniendo en cuenta que las bacterias suelen vivir en medios diluidos, deben realizar un “trabajo” para trasladar muchos de esos nutrientes en contra del gradiente de concentración.
Tradicionalmente se viene considerando tres métodos principales de transporte de sustancias a través de la membrana:
| transporte pasivo inespecífico (= difusión simple); | |
| transporte pasivo específico (= difusión facilitada); | |
| transporte activo. |
Como veremos, los más importantes en procariotas son los sistemas de transporte activo.
2.1 TRANSPORTE PASIVO INESPECÍFICO O DIFUSIÓN SIMPLE
Este transporte consiste en la difusión pasiva de ciertas sustancias para las que la membrana es impermeable, debido a la diferencia de concentración (DC) a ambos lados de dicha membrana (la sustancia tiene mayor concentración fuera que dentro de la célula). Aparte de esta diferencia de concentración, en la difusión pasiva influyen:
| la constante de permeabilidad (P), es decir, el grado de permeabilidad de la membrana a la sustancia en cuestión; | |
| el área o superficie total (A) a través de la que se produce el transporte. |
Las membranas citoplásmicas son impermeables en sí mismas a la mayor parte de las moléculas. Sólo se da en el caso de O2, CO2, NH3, agua y otras pequeñas sustancias polares no ionizadas.
La difusión simple se produce por el paso de estas sustancias a través de poros inespecíficos de la membrana citoplásmica.
2.2 TRANSPORTE PASIVO ESPECÍFICO O DIFUSIÓN FACILITADA
Es un proceso que permite el paso de compuestos por difusión a través de transportadores estereoespecíficos y (al igual que en el caso anterior) sobre la base de un gradiente de concentración (en la dirección termodinámicamente favorable).
El transportador suele ser una proteína integral de membrana (permeasa o facilitador), cuya conformación determina un canal interior, y por el cual un determinado sustrato puede alcanzar el interior, sin gasto de energía. Se piensa que cuando el soluto se une a la parte de la permeasa que da al exterior, esta proteína sufre un cambio conformacional que libera la molécula en el interior. Como al entrar la molécula, enseguida entra en el metabolismo y desaparece como tal, esto basta para mantener el gradiente de concentración que permite esta difusión. La difusión facilitada exhibe propiedades similares a las de las reacciones enzimáticas:
| Especificidad de sustrato: cada permeasa transporte un solo tipo de sustratos químicamente parecidos. | |
| Cinética de saturación de tipo Michaelis-Menten, es decir, la velocidad de transporte aumenta con la concentración de sustrato, hasta un valor límite (Vmax) por encima del cual ulteriores aumentos del soluto no aumentan dicha velocidad (debido a que todas las porinas disponibles están ya totalmente ocupadas): |
Velocidad de entrada: Vent = Vmáx · [Sext] /Km + [Sext]
Velocidad de salida: Vsal = Vmáx · [Sint] /Km + [Sint]
Aunque este sistema de transporte es muy común en eucariotas, es muy raro encontrarlo en bacterias. La explicación evolutiva es que los procariotas suelen vivir en ambientes con pocas concentraciones de nutrientes, y por lo tanto no es frecuente que se den gradientes adecuados. Una de las pocas excepciones la constituye el glicerol, que es transportado por difusión facilitada en una amplia gama de bacterias, tanto Gram-positivas como Gram-negativas. Conforme el glicerol entra, es rápidamente convertido a glicerol-fosfato; por lo tanto, la concentración interna de glicerol como tal es prácticamente nula, lo que facilita esta difusión incluso a bajas concentraciones exteriores de esta sustancia. En Zymomonas existe un facilitador de membrana que transporta glucosa.
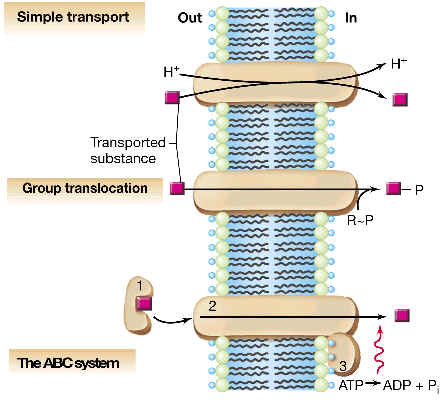
2.3 TRANSPORTE ACTIVO
Consiste en el transporte de sustancias en contra de un gradiente de concentración, lo que requiere un gasto energético. En la mayor parte de los casos este transporte activo (que supone un trabajo osmótico) se realiza
| a expensas de un gradiente de H+ (potencial electroquímico de protones) previamente creado a ambos lados de la membrana, por procesos de respiración y fotosíntesis; | |
| por hidrólisis de ATP. |
Los sistemas de transporte activo son los más abundantes entre las bacterias, y se han seleccionado evolutivamente debido a que en sus medios naturales la mayoría de los procariotas se encuentran de forma permanente o transitoria con una baja concentración de nutrientes.
Los sistemas de transporte activo están basados en permeasas específicas e inducibles. El modo en que se acopla la energía metabólica con el transporte del soluto aún no está dilucidado, pero en general se maneja la hipótesis de que las permeasas, una vez captado el sustrato con gran afinidad, experimentan un cambio conformacional dependiente de energía que les hace perder dicha afinidad, lo que supone la liberación de la sustancia al interior celular.
Estudiaremos los siguientes tipos de transporte activo:
| transporte activo ligado a simporte de protones; | |
| transporte activo ligado a simporte de iones Na+ | |
| transporte activo dirigido por ATP | |
| transporte acoplado a translocación de grupos. |
2.3.1 TRANSPORTE ACTIVO LIGADO A SIMPORTE DE PROTONES
Como se recordará, el simporte se puede definir como el transporte simultáneo de dos sustratos en la misma dirección, por un mismo transportador sencillo. En el caso del transporte activo ligado a simporte de protones, lo que ocurre es que uno de los sustratos (H+) ha creado previamente un gradiente de concentración, cuya disipación es aprovechada por el otro sustrato para entrar con él. Este otro sustrato puede ser:
| una molécula de carga negativa: en este caso, su simporte ligado a protones tiende a disipar sólo el gradiente de concentración. Ejemplos: transporte de iones fosfato, de glutamato, etc. | |
| una molécula neutra: en este caso, su simporte tiende a disipar no sólo el gradiente de concentración, sino también el gradiente eléctrico. Ejemplo: en Escherichia coli, la lactosa usa una ß-galactósido-permeasa, que es una de las permeasas bacterianas más intensamente estudiadas. |
Por otro lado, ciertas moléculas catiónicas (iones K+, lisina), son transportadas directamente a través de permeasas, en ausencia de simporte de protones.
2.3.2 TRANSPORTE ACTIVO LIGADO A SIMPORTE DE IONES SODIO
Se puede considerar una versión modificada del anterior: algunas sustancias no son transportadas activamente de forma directa por el potencial electroquímico de protones, sino indirectamente, a través de un gradiente de Na+ que a su vez se origina a expensas de dicha fuerza protón-motriz (fpm).
El sustrato entra por una permeasa, junto con iones Na+, pero a su vez este sodio se recicla por un sistema de antiporte, a expensas de la disipación del potencial de protones.
Ejemplo: el azúcar melibiosa, en el caso de la enterobacteria E. coli.
Estos dos tipos de transporte activo ligados a simporte quedan inhibidos si tratamos las células con algún agente ionóforo (p. ej., el antibiótico valinomicina), que destruye el potencial electroquímico de protones.
El transporte activo ligado a simporte de iones (H+, Na+) resulta muy económico, ya que sólo se gasta un protón por cada molécula transportada, mientras que por cada ATP sintetizado se suelen gastar 3 protones que se disipan en las ATPasas. Las permeasas que realizan este transporte suelen ser proteínas integrales de membrana provistas de unos 12 segmentos transmembranosos en configuración de a-hélice.
2.3.3 TRANSPORTE ACTIVO DIRIGIDO POR ATP
El tipo paradigmático de este tipo de transporte se denomina de transportadores ABC o ATPasas de tráfico, y se conocen muchos ejemplos en eubacterias y arqueas. Vamos a describir el caso de un sistema ABC en enterobacterias (como E. coli). Se trata de un sistema de varios componentes, en el que existen proteínas periplásmicas que captan el sustrato con gran afinidad, y lo llevan hasta unas proteínas de membrana, las cuales acoplan el paso de dicho sustrato hasta el citoplasma (sin alteralo químicamente) con la hidrólisis de ATP
http://www.ugr.es/~eianez/Microbiologia/06membrana.htm



No hay comentarios:
Publicar un comentario